Volúmenes Respiratorios
En reposo, un adulto promedia 18 (mujer) y 16 (hombre) respiraciones por minuto, en los que entran y salen de los pulmones casi 500 mililitros de aire.
Se pueden distinguir 4 volúmenes pulmonares que sumados, son iguales al máximo volúmen al que es posible expandir los pulmones:
1- El volúmen corriente es el volúmen de aire inspirado o espirado en cada respiración normal, es de aproximadamente 500 mililitros.
2- El volúmen de reserva inspiratorio es el volúmen adicional de aire que se puede inspirar por encima del volúmen corriente normal, es de aproximadamente 3000 mililitros.
3- El volúmen de reserva espiratorio es la cantidad adicional de aire que se puede espirar por espiración forzada después de una espiración normal, es de aproximadamente 1100 mililitros.
4- El volúmen residual es el volúmen de aire que queda en los pulmones tras la espiración forzada, es de aproximadamente 1200 mililitros.
La suma del volúmen de reserva inspiratorio, más el volúmen corriente, más el volúmen de reserva espiratorio se conoce como capacidad vital y es la máxima cantidad de aire que puede expulsar una persona de los pulmones después de una inspiración máxima y espirando al máximo. Es de unos 4600 mililitros.
Todos los volúmenes pulmonares y la capacidad vital pulmonar son entre un 20 y 25% menores en la mujer que en el hombre, y son mayores en personas altas y atléticas que en personas pequeñas y que no realizan ejercicio.
Si durante la respiración tranquila se intercambian 500 mililitros de aire con el exterior, en un minuto ese volúmen llega a 7 u 8 litros. Si la frecuencia respiratoria aumenta, ese volúmen se hace mucho mayor, como ocurre durante el ejercicio físico, donde el volúmen de aire intercambiado llega a 100-120 litros por minuto.
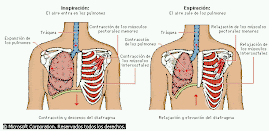
Es decir que, para aumentar la ventilación pulmonar el organismo cuenta con tres mecanismos: 1- aumento de la amplitud de los movimientos respiratorios, 2- aumento de su frecuencia, 3- a largo plazo, un aumento del volúmen de la caja toráxica y de los pulmones.
Un método simple de estudiar la ventilación pulmonar es registrar la variación del volúmen de aire que entra y sale de los pulmones, un proceso denominado espirometría. El espirómetro, consiste en un tambor invertido sobre una cámara de agua, con el tambor contrapesado por un peso. En el tambor existe un gas respirable, aire u oxígeno; un tubo conecta la boca con la cámara del gas.
Cuando se respira a la cámara o se toma aire de ella, el tambor sube y baja y se hace un registro adecuado sobre una hoja de papel en movimiento.
La ventilación alveolar: lógicamente, la parte más importante de todo el proceso mecánico de la respiración, es la ventilación de los alvéolos pulmonares. Pero ¿todo el aire que el aparato respiratorio intercambia con el exterior llega a los alvéolos? La respuesta es negativa porque una parte del aire que ingresa debe, necesariamente, rellenar las fosas nasales, la faringe, la laringe, la tráquea, y los bronquios; en ninguno de esos órganos se cumple la función respiratoria propiamente dicha. El volúmen de aire que albergan esos órganos llega a unos 150 mililitros, por el motivo antes mencionado se le denomina espacio muerto anatómico. Todavía, el aire llega a muchos alvéolos en los que, en determinados momentos, no llega sangre para efectuar intercambio de gases. El aire que rellena esos alvéolos sumado al del espacio muerto anatómico constituye el espacio muerto fisiológico.